Conspect al capitolului ”The Eye”, de Bear, Connors și Paradiso, în ”Neuroscience. Exploring the Brain”, în 2016.
Introducere
Lumina este energie electromagnetică emisă sub formă de unde. Aceste unde se lovesc de obiecte și astfel sunt absorbite, împrăștiate, reflectate sau curbate. Din aceste interacțiuni, sistemul vizual poate extrage informații despre lumea înconjurătoare.
Sistemul vizual a oferit noi modalități de comunicare, de predicție a traiectoriei obiectelor și a evenimentelor în timp și spațiu, dar a permis și apariția unor noi forme de imagerie mentală și abstractizare.
Peste o treime din cortexul cerebral este implicat în analizarea lumii vizuale.
Sistemul vizual al mamiferelor începe cu ochiul, iar în ochi se află retina, ce conține fotoreceptori specializați pentru convertirea energiei luminoase în activitate neuronală. Restul ochiului funcționează ca o cameră foto. Însă retina nu înregistrează doar pasiv niveluri diferite de lumină. Outputul retinei nu este o reproducere fidelă a intensității luminii care cade pe ea. Retina este mai degrabă specializată în detectarea diferențelor de intensitate a luminii ce cade pe diferite părți ale ei.
Axonii neuronilor retinieni sunt adunați în nervi optici, care distribuie informația sub forma potențialelor de acțiune către mai multe structuri cerebrale, inclusiv cele care se ocupă de reglarea ritmurilor biologice și celor care controlează poziția ochilor și elementele de optică. Primul releu sinaptic îl fac nervii optici în nucleul geniculat lateral din talamusul dorsal. De aici, semnalele sunt transmise către cortexul cerebral.
Proprietățile luminii
Lumina
Lumina este radiație electromagnetică vizibilă ochilor noștri. Această radiație ia forma unei unde de energie. Undele sunt caracterizate de lungime, frecvență și amplitudine.
Lungimea de undă este distanța dintre vârfuri succesive. Frecvența este numărul de unde pe secundă, iar amplitudinea este diferența dintre baza și vârful undei.
Conținutul de energie din radiația electromagnetică este proporțional cu frecvența ei. Radiația emisă cu o frecvență ridicată are conținut energetic mai mare.
Lumina vizibilă are lungimi de undă între 400 și 700 nm. Combinația de lungimi de undă din acest interval, emisă de soare, este percepută de ochiul uman ca fiind albă, în timp ce o singură lungime de undă apare ca fiind una dintre culorile curcubeului.
Optica
În vid, lumina circulă în linie dreaptă și poate fi descrisă drept rază. O rază de lumină în mediul nostru se deplasează tot în linie dreaptă, însă se lovește de atomii și moleculele din atmosferă și din obiecte. Aceste interacțiuni includ reflexia, absorbția și refracția.
Reflexia este respingerea luminii de către o suprafață. Ea are unghiuri diferite, în funcție de unghiul la care lumina a lovit suprafața respectivă.
Absorbția este transferul de energie către suprafața de care se ciocnește lumina. Suprafețele care par negre absorb energia tuturor lungimilor de undă vizibile, pe când alte substanțe absorb o parte din lungimile de undă și reflectă altele. Acesta este principiul care stă la baza pigmenților din vopsele.
Refracția este îndoirea razelor de lumină ce are loc atunci când acestea trec dintr-un mediu transparent în altul. Cel mai evident exemplu este atunci când lumina trece din aer în apă.
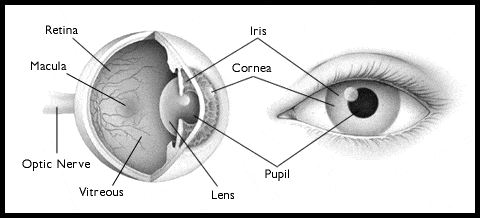
Ochiul: structura
Anatomia grosieră a ochiului
Pupila este deschiderea ce permite luminii să intre în ochi și să ajungă la retină. Pare neagră datorită pigmenților ce absorb lumina și care se găsesc pe retină. Pupila este înconjurată de iris. Acesta conține doi mușchi ce pot modifica dimensiunea pupilei. Unul o face mai mică atunci când se contractă, altul o face mai mare când se contractă. Pupila și irisul sunt acoperite de cornee, o suprafață sticloasă, transparentă. Corneea se continuă cu sclera, care este ”albul ochilor”.
Globul ocular este poziționat în interiorul scalpului, în orbita oculară și este fixat de trei perechi de de mușchi extraoculari, atașați de scleră. Acești mușchi nu sunt vizibili, fiind acoperiți de conjunctivă, o membrană care conectează interiorul pleoapelor cu sclera.
Nervul optic ce conține axonii celulelor retiniene este conectat de globul ocular în partea posterioară a acestuia.
Structura oftalmoscopică a ochiului
Retina poate fi analizată la oftalmoscop. Primele elemente vizibile sunt vasele de sânge de pe suprafața ei. Acestea pornesc din discul optic, locația prin care ies și fibrele nervoase din retină. Această arie nu analizează lumina, la fel cum nu poate fi analizată lumina nici pe ariile acoperite de vase de sânge. Aceste ”pete oarbe” sunt compensate la nivel perceptiv de creier.
În mijlocul retinei se află macula, partea din retină alocată vederii centrale (spre deosebire de cea periferică).
O altă structură este fovea, care apare la oftalmoscop ca un punct negru de aproximativ 2 mm. Aici este retina cea mai subțire.
Foveeea se află exact în centrul retinei, funcționând și ca punct de referință. Partea din retină de la fovee înspre nas se numește nazală, iar cea înspre tâmplă se numește retină temporală. Deasupra foveei este retina superioară, iar sub fovee cea inferioară.
Secțiunea transversală a ochiului
Dinspre exterior, primul strat este reprezentat de cornee. În spatele ei se află umoarea apoasă, care are și rol de hrănire a corneei. Urmează irisul, iar în spatele acestuia lentila transparentă. Aceasta este suspendată de ligamente atașate mușchiului ciliar, ce formează un cerc în interiorul ochiului.
Lentila împarte interiorul ochiului în două compartimente umplute de fluide diferite. Umoarea apoasă este între cornee și lentilă, iar umoarea vitroasă, care are o consistență gelatinoasă, se găsește între lentilă și retină. Are rolul de a păstra forma sferică a ochiului.
Formarea imaginii de către ochi
Aducerea obiectelor în focus este realizată prin capacitatea de refracție a corneei și lentilei. Corneea are cea mai mare putere de refracție pentru că lumina ajunge la ochi din aer, iar corneea este compusă în principal din apă. Refracția se datorează încetinirii luminii odată ce ajunge în apă, comparativ cu viteza pe care o are în aer.
Refracția de către cornee
Distanța de la suprafața refractară la punctul în care converg razele paralele de lumină se numește distanța focală. Aceasta depinde de curbura corneei: cu cât este mai curbată, cu atât este mai scurtă distanța focală. Reciproca distanței focale în metri este o unitate de măsură numită dioptrie. Corneea are o putere de refracție de aproximativ 42 de dioptrii, ceea ce înseamnă că razele paralele care ating suprafața corneei vor converge la 2,4 cm în spatele ei. Aceasta este distanța aproximativă dintre cornee și retină. Vezi imaginea de la pagina 301.
Acomodarea de către lentilă
Și lentila contribuie cu aproximativ 12 dioptrii la formarea unei imagini clare a unui punct îndepărtat. Însă lentila este mai importantă pentru formarea imaginilor clare ale obiectelor aflate mai aproape de 9 m de la ochi.
Când obiectele sunt apropiate, razele de lumină nu mai sunt paralele. Prin urmare, este nevoie de o putere de refracție mai mare pentru a le reprezenta clar pe retină. Această putere suplimentară este oferită de acomodarea lentilei. Acest proces presupune contractarea mușchiului ciliar care reduce spațiul inelului interior. Astfel, lentila devine mai rotundă și mai groasă, crescând curbura suprafețelor acesteia și crescându-i, implicit, puterea de refracție.
Reflexul pupilar la lumină
Pupila contribuie la funcționarea optică a ochiului prin ajustarea continuă la nivelurile diferite de lumină ambientală. Reflexul pupilar la lumină se datorează unor conexiuni între retină și neuroni ai trunchiului cerebral ce controlează mușchii ce modifică dimensiunea pupilei.
Acest reflex este consensual: dacă un singur ochi este expus la o lumină mai puternică, ambii ochi reacționează prin constricție pupilară. Absența acestei consensualități este considerată semn al unei afectări grave la nivelul trunchiului cerebral.
Un efect benefic al constricției pupilare este o îmbunătățire a profunzimii percepțieie, adică a distanței la care poate fi un obiect aflat în focus.
Câmpul vizual
Câmpul vizual este spațiul ce poate fi văzut de retină atunci când ochiul este fixat drept înainte. Acesta se extinde aproximativ 100 de grade în partea temporală și 60 de grade în partea nazală a retinei, unde vederea este blocată de nas.
Partea stângă a câmpului vizual este proiectată în partea dreaptă a retinei, iar partea dreaptă a câmpului vizual este proiectată în partea stângă a retinei. La fel, partea superioară a câmpului vizual este proiectată în partea inferioară a retinei și viceversa.
Acuitatea vizuală
Abilitatea ochiului de a distinge două puncte apropiate unul de celălalt este numită acuitate vizuală. Aceasta depinde de spațierea fotoreceptorilor de pe retină și de precizia refracției ochiului.
Distanța de-a lungul retinei poate fi descrisă în termeni de grade ale unghiului vizual.
Putem vorbi de abilitatea ochiului de a diferenția puncte care sunt separate de un anumit număr de grade ale unghiului vizual. Panoul Snellen este cel pe care îl citim la oftalmolog pentru a ne evalua acuitatea. O acuitate perfectă presupune recunoașterea unor litere care se întind pe mai puțin de un grad al unghiului vizual (0,083 grade).
Anatomia microscopică a retinei
Sub influența luminii, fotoreceptorii influențează potențialul membranar al celulelor bipolare cu care sunt conectați. La rândul lor, celulele bipolare transmit semnalul mai departe celulelor gliale care descarcă potențiale de acțiune prin nervul optic înspre creier.
Celulele orizontale primesc și ele semnale de la fotoreceptori și influențează, la rândul lor, celulele bipolare și fotoreceptorii din preajmă. O gamă largă de celule amacrine primesc semnale de la celule bipolare și se conectează în lateral, influențând celulele ganglionare, cele bipolare și alte celule amacrine dimprejur.
Singurele celule fotosensibile din retină sunt fotoreceptorii cu formă de conuri sau de bastonașe. Singurul output al retinei sunt celulele gliale. Doar ele descarcă potențiale de acțiune.
Organizarea laminară a retinei
Organizarea laminară a retinei presupune existența mai multor straturi ce par aranjate din afară înspre înăuntru. Lumina trece prin umoarea vitroasă, pe lângă celulele ganglionare și bipolare, care sunt relativ transparente, înainte de a ajunge la fotoreceptorii de pe retină.
Epiteliul pigmentat din spatele fotoreceptorilor absoarbe lumina ce trece prin retină, minimizând împrăștierea luminii în ochi astfel încât să afecteze calitatea imaginii formate.
Stratul celulelor ganglionare conține corpurile acestor celule. Mergând înspre exteriorul ochiului, urmează stratul nuclear interior, format din corpurile celulelor bipolare, orizontale și amacrine, iar apoi stratul nuclear exterior, format din corpurile celulare ale fotoreceptorilor.
Între stratul celulelor ganglionare și cel nuclear interior se află un strat plexiform interior care conțină sinapsele dintre celulele bipolare, amacrine și ganglionare.
Între straturile nucleare interior și exterior se află stratul plexiform extern ce conține sinapsele fotoreceptorilor cu celulele bipolare și orizontale.
Un ultim strat este cel al segmentelor exterioare ale fotoreceptorului. Acesta conține elementele foto-sensibile ale retinei. Aceste segmente exterioare sunt inserate în epiteliul pigmentat.
Structură fotoreceptori
Fiecare fotoreceptor are patru regiuni: segmentul extern, segmentul intern, corpul celular și terminația sinaptică.
Segmentul extern conține discuri membranoase cu fotopigmenți sensibili la lumină. Acești pigmenți absorb lumina și determină modificări ale potențialului membranei fotoreceptorului.
Există două tipuri de fotoreceptori: bastonașele și conurile. Primele au un segment extern lung, cilindric, ce conține multe discuri. Conurile au un segment extern mai scurt , de formă conică și cu mai puține discuri membranoase. Datorită diferenței în concentrația discurilor și fotopigmenților, bastonașele sunt de 1000 de ori mai sensibile la lumină, comparativ cu conurile. Vezi pagina 307 pentru o ilustrare grafică a diferențelor.
În condiții de obscuritate (scotopice), doar bastonașele contribuie la formarea imaginii retiniene. În schimb, în condiții de luminozitate bună (fotopice), conurile au cea mai mare contribuție. În condiții intermediare (mezopice), atât conurile cât și bastonașele contribuie la formarea imaginii retiniene.
Toate bastonașele conțin același fotopigment, dar există trei tipuri de conuri, fiecare cu câte un pigment diferit. Aceste diferențe determină responsivitatea la lungimi de undă diferite ale luminii pentru aceste tipuri de conuri. În plus, doar conurile ne ajută să percepem culoarea, nu și bastonașele.
Diferențe regionale în structura retinei și consecințele lor vizuale
Majoritatea conurilor se găsesc în fovee, iar proporția lor scade pe parcurs ce ne apropiem de periferie. Bastonașele sunt însă absente din fovee, dar se gâsesc într-o proporție mai mare la periferie.
Atunci când lumina este la niveluri fopopice (lumina zilei), avem o sensibilitate spațială mai mare în centrul retinei.
De asemenea, suntem mai puțin capabili să diferențiem culorile în ariile periferice ale retinei datorită numărului redus de conuri care se găsesc în aceste arii.
Suntem mai sensibili la niveluri scotopice de lumină (luminozitate redusă) în zonele periferice ale retinei, deoarece bastonașele răspund mai puternic la acest tip de lumină decât conurile, iar aceste bastonașe sunt mai multe la periferia retinei și lipsesc din fovee.
Nu suntem capabili să diferențiem culorile noaptea deoarece percepția culorii este apanajul exclusiv al conurilor.
Fototransducția
Fototransducția se referă la transformarea energiei luminii în modificări ale potențialului membranei.
Fototransducția în bastonașe
O modalitate de a reprezenta informația în sistemul nervos este prin modificările ce au loc la nivelul potențialului membranei neuronale.
Stimularea fotopigmentului de către lumină activează proteinele G, care activează o enzimă efectoare ce schimbă concentrația citoplasmatică a unei molecule mesager secundare. Această modificare determină închiderea unui canal ionic, schimbând astfel potențialul membranei.
Potențialul membranei neuronale este de în condiții normale de -65 mV (microvolți), iar în condiții de întuneric complet acest potențial este de -30 mV. Această depolarizare are loc datorită influxului ionilor de sodiu prin canale speciale din segmentul extern al membranei. Această trecere al încărcării pozitive prin membrană în condiții de întuneric se numește curent întunecat (dark current).
Canalele de sodiu sunt stimulate să se deschidă de către un mesager intracelular secundar numit monofosfat- guanozină ciclică (cyclic guanosine monophosphate sau cGMP). Acest mesager este produs în fotoreceptor de enzima guanilil-ciclază.
Lumina reduce cGMP, determinând canalele de sodiu să se închidă, iar potențialul membranei devine mai negativ.
Prin urmare, reacția fotoreceptorilor la lumină este una de hiperpolarizare.
Acesta este inițiat prin absorbirea radiației electromagnetice de către un fotopigment din membrană. Acest fotopigment se numește rodopsină și este o proteină receptoare care are un agonist asociat, numit retinal (un derivat al vitaminei A). Absorbirea luminii modifică retinalul astfel încât să activeze opsina.
Acest proces este cunoscut drept albire deoarece fotopigmentul își schimbă efectiv culoarea din violet în galben. Albirea rodopsinei stimulează proteina G numită transducină din membrana discului, care în schimb activează enzima efectoare fosfodiesterază (PDE). Aceasta descompune cGMP, care este prezentă în mod normal în citoplasma bastonașului (când e întuneric).
Reducerea cGMP determină canalele ionilor de sodiu să se închidă, iar membrana să se hiperpolarizeze.
O consecință funcțională a acestui întreg proces este amplificarea semnalului. Mai multe proteine G sunt activate de fiecare moleculă de fotopigment și fiecare enzimă PDE descompune mai multe molecule cGMP. Această amplificare conferă o sensibilitate foarte mare a sistemului vizual la intensități reduse de lumină.
Bastonașele sunt mai sensibile la lumină comparativ cu conurile pentru că ele conțin mai multe discuri pe segmentele exterioare și, implicit, au mai mult fotopigment. De asemenea, ele amplifică răspunsul la lumină mai mult decât conurile.
Pe scurt, etapele transducției luminii în bastonașe sunt:
- Lumina activează (albește) rodopsina.
- Este stimulată transducina (o proteină G).
- Este activată fosfodiesteraza (PDE, o enzimă efectoare).
- Activitatea PDE reduce nivelul cGMP.
- Canalele de sodiu se închid, iar membrana celulară se hiperpolarizează.
Fototransducția în conuri
Vederea în timpul zilei depinde exclusiv de activitatea conurilor. Acestea au fotopigmenți ce necesită mai multă energie pentru a fi albiți.
Procesul de fototransducție în conuri este analog celui din bastonașe. Diferența este dată de tipurile de opsine din discurile membranare ale conurilor. Aceste opsine din conuri sunt de trei feluri, fiecare sensibilă la anumite intervale ale lungimii de undă. Conurile pentru lungime de undă mică sunt numite albastre, cele pentru lungime de undă medie sunt numite verzi, iar cele pentru lungime de undă mare sunt numite roșii. Ele răspund maximal la lumina cu lungimea de undă de 430, 530, respectiv 560 nm.
Perceperea culorii
Culoarea pe care o percepem este determinată de contribuția relativă la semnalul retinei a conurilor pentru lungimi de undă scurtă, medie și lungă.
Teoria tricromatică Young-Helmholtz susține că creierul alocă o culoarea unui obiect pe baza outputului comparativ al celor trei tipuri de conuri. Când toate cele trei sunt la fel de active, atunci percepem ”alb”.
Adaptarea la întuneric și la lumină
Trecerea de la vederea bazată exclusiv pe activitatea conurilor din timpul zilei la cea bazată exclusiv pe bastonașe în timpul nopții nu are loc instantaneu. În funcție de intensitatea luminii inițiale, poate dura de la câteva minute la aproape o oră pentru a obține cel mai înalt nivel de sensibilitate în întuneric. Acest fenomen se numește adaptarea la întuneric. Prin acest proces, sensiblitatea la lumină se îmbunătățește de un milion de ori sau chiar mai mult.
Această adaptare este explicată în primul rând de modificarea diametrului pupilar, care permite unei cantități mai mari de lumină să intre. Însă aceasta nu crește sensiblitatea la lumină decât cu un factor de 10.
Componenta mai importantă a adaptării la întuneric este regenerarea rodopsinei nealbite și o ajustare a circuitelor funcționale ale retinei astfel încât informația de la mai multe bastonașe este disponibilă pentru fiecare celulă ganglionară.
Datorită acestei creșteri masive a sensibilității la lumină, atunci când procesul este inversat, iar ochiul adaptat la întuneric este expus la lumină puternică, apare o saturație temporară. În primele 5-10 minute are loc adaptarea la lumină, inversând procesele mai sus prezentate.
Rolul calciului în adaptarea la lumină
Dincolo de factorii deja prezentați, abilitatea ochiului de a se adapta la variațiile de intensitate a luminii se datorează și modificărilor de concentrație a calciului din conuri.
Canalele de sodiu cGMP-dependente permit și trecerea particulelor de calciu. În întuneric, ionii de calciu intră în conuri și inhibă efectul enzimei ce sintetizează cGMP. Odată închise canalele cGMP-dependente, fluxul ionilor de calciu în interiorul fotoreceptorului este oprit, odată cu fluxul ionilor de sodiu. Prin urmare, cGMP este sintetizat, permițând canalelor cGMP-dependente să se redeschidă.
Adaptarea locală la întuneric, lumină și culoare
Albirea fotoreceptorilor și alte mecanisme adaptative, cum ar fi influența calciului asupra cGMP are loc nu global, ci și la nivelul fiecărui con în parte.
Procesarea și outputul retinian
Doar celulele ganglionare transmit potențiale de acțiune. Toate celelalte celule din retină (cu excepția unor celule amacrine) răspund la stimulare prin modificări graduale ale potențialului membranar.
Fotoreceptorii secretă neurotransmițători (amino acid glutamat) când sunt depolarizați. Depolarizarea are loc la întuneric, iar la lumină se fotoreceptorii se hiperpolarizează.
În stratul plexiform extern, fiecare fotoreceptor face sinapse cu celule bipolare și celule orizontale. Cele bipolare leagă fotoreceptorii de celulele ganglionare. În schimb, celulele orizontale transmit informația lateral pentru a influența alte celule bipolare și fotoreceptori din apropiere.
Câmpul receptor
Suprafața retinei care modifică rata de descărcare a unui neuron atunci când este proiectată lumina pe această suprafață se numește câmp receptor. Dacă lumina este proiectată în afara acestui câmp, ea nu va influența activitatea neuronului respectiv.
La nivelul retinei, pata (spot) de lumină oferă răspunsurile optime de la celulele ganglionare, dar în arii diverse ale cortexului vizual, neuronii răspund cel mai bine la linii luminoase sau chiar forme complexe cu semnificați biologică, cum sunte fețele sau mâinile.
Câmpurile receptoare ale celulelor bipolare
Celulele bipolare pot fi împărțite în două tipuri: ON și OFF, în funcție de răspunsul la glutamatul eliberat de fotoreceptori.
Celulele bipolare OFF sunt cele care se hiperpolarizează când lumina interacționează cu un con. Invers, când interacțiunea luminii cu un con depolarizează celula bipolară, aceasta este celulă ON.
Celulele bipolare OFF au receptori de glutamat ionotropici, iar aceste canale glutamat-dependente mediază depolarizarea excitatorie. Hiperpolarizarea conului determină secretarea unei cantități mai reduse de neurotransmițător, rezultând într-o mai mare hiperpolarizare a celulei.
Pe de altă parte, celulele bipolare ON au receptori metabotropici și răspund la glutamat prin hiperpolarizare.
Fiecare celulă bipolară primește input sinaptic direct de la un grup de fotoreceptori. Numărul acestora variază de la 1 în centrul foveei și până la mii în zona periferică a retinei.
Celulele bipolare sunt conectate și prin celule orizontale la un inel de fotoreceptori care înconjoară clusterul central.
Când un fotoreceptor hiperpolarizează ca răspuns la lumină, outputul este trimis celulelor orizontale care hiperpolarizează și ele. Efectul acestei hiperpolarizări a celulelor orizontale este de a contracara efectul luminii asupra fotoreceptorilor din vecinătate.
Pe scurt, câmpul receptor al unei celule bipolare are două părți: o zonă circulară care conferă inputu fotoreceptor direct și care se numește centrul câmpului receptor și o arie înconjurătoare care oferă input prin celulele orizontale și care se numește încercuire a câmpului receptor (receptive field surroud). Răspunsul potențialului membranar al unei celule bipolare la lumină în centrul câmpului este opus celui la lumină în jurul câmpului. Se spune că aceste celule au câmpuri receptoare centru-încercuire antagoniste.
Organizarea câmpului receptor centru-încercuire este transferată de la celulele bipolare la cele ganglionare prin sinapsele din stratul plexiform interior. Conexiunile laterale ale celulelor amacrine din stratul plexiform interior contribuie și ele la elaborarea câmpurilor receptoare ale celulei ganglionare și la integrarea inputului de la conuri și de la bastonașe în celulele ganglionare.
Câmpurile receptoare ale celulei ganglionare
Cele mai multe celule ganglionare au același câmp receptor concentric, cu centru și periferie, analog celulelor bipolare. Diferența este că celulele ganglionare descarcă potențiale de acțiune.
O celulă ganglionară ON va fi depolarizată și va emite o salvă de potențiale de acțiune când un mic punct de lumină este proiectat pe centrul câmpului său receptor. În schimb, celula ganglionară OFF va descărca mai puține potențiale de acțiune când un punct mic de lumină este proiectat în centrul câmpului său receptor. Acest tip de celulă va descărca mai multe potențiale când pe centrul său receptor este proiectat un punct întunecat
Cele mai multe celule ganglionare par să fie responsive la diferențe de iluminare ce au loc în câmpurile lor receptoare.
Organizarea centru-periferie a câmpurilor receptoare determină o accentuare a contrastului muchiilor dintre suprafețe iluminate și suprafețe întunecate.
Relațiile dintre structură și funcție
Se face o diferențiere între celulele ganglionare de tip M, care sunt mai mari și reprezintă doar 5% dintre celulele ganglionare, și celulele ganglionare de tip P, care sunt mai mici și reprezintă 90% dintre celulele ganglionare. Restul de 5% sunt numite celule ganglionare nonM-nonP.
Celulele M au câmpuri receptoare mai mari, conduc potențialele de activare mai rapid prin nervul optic și sunt mai sensibile la stimulii cu contrast redus. Ele răspund la stimulare în centrele câmpurilor receptoare printr-o explozie temporară a potențialului de acțiune, pe când celulele P răspund printr-o descărcare susținută pe tot parcursul prezenței stimulului.
Celulele ganglionare culorilor opuse
Unele celule P și non-M-nonP sunt sensibile la diferențele dintre lungimile de undă ale luminii. Majoritatea acestor neuroni sensibili la culoare sunt numiți celule ale culorilor opuse (color-opponent cells) pentru că răspunsul la o culoare din centrul câmpului receptor este anulată prin expunerea unei alte culor în încercuirea câmpului receptor.
Într-o celulă cu un centru ON roșu și o periferie (surround) OFF verde, centrul câmpului receptor este alimentat de conuri roșii, iar încercuirea este alimentată de conuri verzi printr-un circuit inhibitor de celule orizontale.
Dacă pe centrul câmpului receptor cade lumină roșie, neuronul răspunde cu o izbucnire a potențialelor de acțiune. Dacă lumina roșie acoperă și centrul și periferia, neuronul este în continuare activat, dar într-o mai mică măsură. Lumina roșie activează și periferia OFF verde pentru că lungimile de undă roșii ale luminii sunt parțial absorbite de conurile verzi, iar activarea lor inhibă răspunsul neuronului.
Fotoreceptorii celulei ganglionare
Există un un număr foarte restrâns de celule ganglionare retiniene care au proprietăți fotoreceptoare. Acestea sunt numite intrinsec fotosensibile (intrinsically photosensitive retinal ganglion cells sau ipRGC) și folosesc melanopsina ca fotopigment.
ipRGC funcționează și ca celule ganglionare normale, primit semnale de la conuri și bastonașe. Trimit axoni prin nervul optic, dar sunt și fotoreceptori care se depolarizează când sunt expuși la lumină.
Au un câmp dendritic de mari dimensiuni, iar aceste dendrite sunt fotosensibile, ceea ce înseamnă că aceste celule însumează inputul luminos din zone retiniene mai întinse comparative cu bastonașele sau conurile.
O funcție importantă a ipRGC este de a oferi input ariilor vizuale subcorticale care sincronizează comportamentul la modificările zilnice ale luminozității (ritmurile circadiene).
Procesarea paralelă
Procesarea paralelă în sistemul vizual se referă la faptul că atribute vizuale diferite sunt procesate simultan, folosind căi distincte. Spre exemplu, vedem lumea cu doi ochi ce oferă două fluxuri diferite de informație. Un alt exemplu este dat de fluxurile independente de informație despre lumină și întuneric ce vin de la celulele ganglionare cu centru ON și cele cu centru OFF de pe fiecare retină.
Radu este psiholog, doctor în psihologie, consultant organizațional, antreprenor și editorul fondator al Psihoteca.